レクチャーに使用されたパワーポイントも掲載していますので合わせてご覧ください。
non-coding RNAとは?
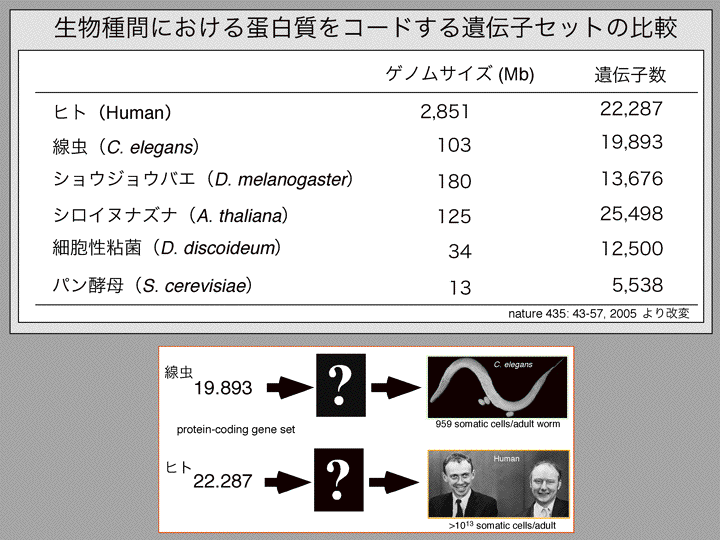
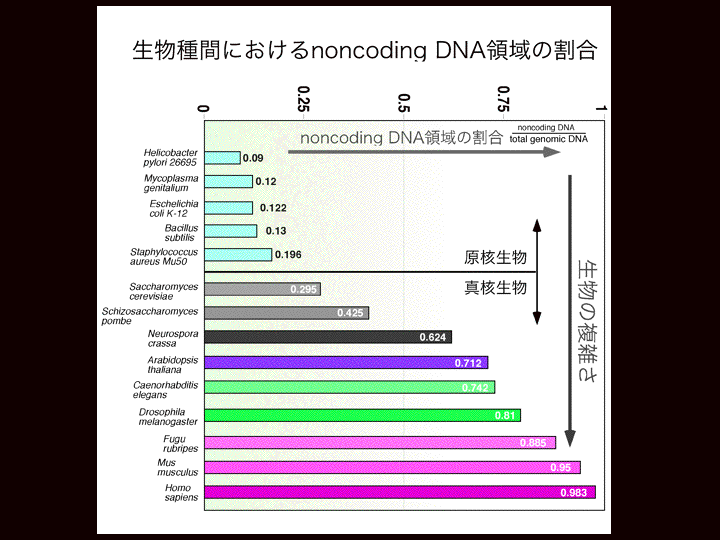
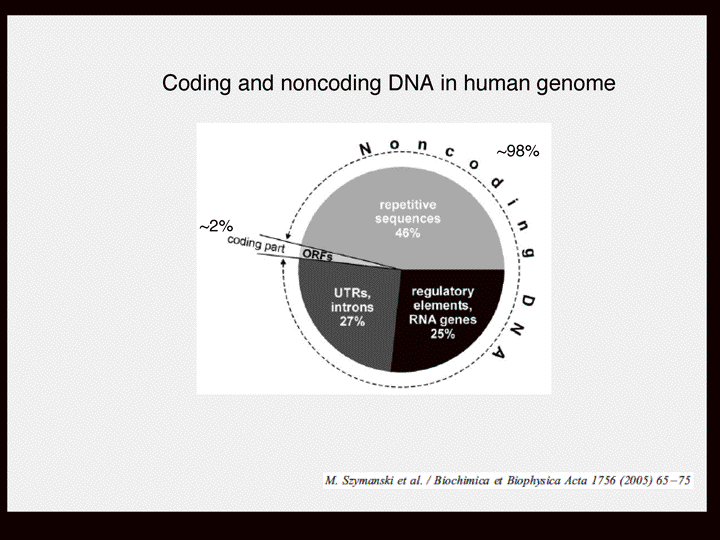
| 塩見 | イントロダクションから始めたいと思います。今、立花さんがおっしゃられたように今何がおこっているかを説明したいと思います。ゲノム解析がここ10年間進んできてまして、300以上の生物種の全ゲノム配列が完了しています。そして現在1000以上の生物のゲノムの解析が進んでいます。 そういうことによって何がわかってきたか。 スライド1 遺伝子数ってものが見えてきました。この場合、遺伝子数というのはタンパク質をコードする数のことです。例えばヒトの場合は二万二千ぐらい、線虫の場合が一万九千、ショウジョウバエの場合が一万三千、それから、植物の遺伝学でよく使われるシロイヌナズナは二万五千ぐらい。これらをみると生物の複雑さと遺伝子の数は相関していない。なぜだろう? 例えば、細胞の数を比べると線虫が千個であるのに対してヒトは精子とかを除いて十の13乗個ぐらいの細胞から成り立っている。ヒトっていうのは非常に複雑で高性能な生き物で、線虫はたしかに高性能ですがそんなに複雑ではありません。でも遺伝子をみてもそんな何オーダーの違いがあるわけではありません。 じゃ、何によって違うのか?そのヒントがこれだと思います。 スライド2 これもゲノム解析の結果わかってきたことですが、DNAをタンパク質をコードする領域とコードしない領域と分けていってその比をとっていく。つまりゲノム全体の中でタンパク質をコードしない領域の割合をプロットしていきますと、例えば(スライド)こちらに大腸菌があります、そして原核生物と真核生物、単構造と分裂構造。こういう風にノンコーディングのDNA(以後ncDNA)の割合は生物が複雑になるほど大きくなって、ヒトで最大になります。 スライド3 ヒトの場合、98%以上がタンパク質をコードしていない領域である。では98%が何をしているかというと一昔前まではジャンクと言われて、つまりゴミだとかカスだとかいうようなものだろうと。何もやっていない、機能していないと思われていた。これがヒトのゲノムを単純に分けた場合どんな風になるか。さきほども言いましたように98%がタンパク質をコードしない領域、タンパク質をコードしている領域がわずか2%ぐらい。そしてその98%のうち半分ぐらいがトランスポゾンと言われるリブ配列からなっていて、その他にイントロンやチフスエレメントなど、遺伝子の発現をコントロールするいろいろなエレメントがあります。特に何もやってない、ヒトに対して悪さをしているのではないかという領域は、最近そうでないということが分かってきた。それはあとで話します。 そしてもう一つ、理研のグループの大きな成果なのですが、最近のゲノム研究で分かってきたことは、転写産物を網羅的に解析していくと、タンパク質をコードする領域は2%なんですが、転写する領域はゲノムの大半、70%で、つまりほとんどがRNAになっているわけです スライド4 じゃ、RNAになって何をしているか? しかも、彼らのグループがやっているのはmRNAの形をとっているものを解析していて、mRNAの形をとっているにも関わらずタンパク質をコードしないもの、いわゆるオープンリーディングフレームが取れないというものは全体の50%以上であることがわかった。 しかも、面白いのはDNAというのは二本鎖ですから上側が読まれる場合と下側が読まれる場合があって、転写されるほうを便宜的にセンスと言い、もう片方をアンチセンスと言うわけです。転写の方向って言うのは右向きと左向きとがあって、センスとアンチセンスという形になるのですが、同じ領域で右向きの転写と左向きの転写が起こっている領域が全体の72%以上あることがわかった。 こういうような何もタンパク質をコードしない部分も含めてゲノムのほとんどが、すくなくとも70%転写されていて、ノンコーディングタイプのmRNA が大量に、つまりmRNA大陸のRNAが大量につくられていてしかもペアで存在することが多い。こういうことが最近のゲノム解析の結果わかった。 タンパク質をコードする部分は非常に少ないけどゲノムの大半は転写されるているんだ、RNAになっているんだ。 |
ncRNA=生物の複雑さ
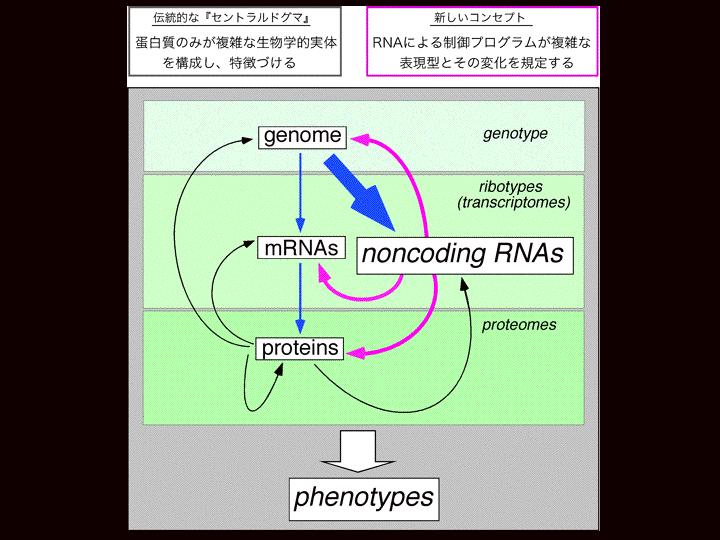
| 塩見 | そのうちタンパク質をコードしない大量のRNAが何をしているのか?っていうのがこれからわたしたちが解決しなければいけない、非常に重要な問題になっていくと思います。 で、その問題の解決が、まさに、”少ないタンパク質をコードする遺伝子を使い回すことによってなぜヒトという複雑な生き物ができるのか?”という問いに答えることになると思います。 スライド5 いままでのセントラルドグマっていうのはゲノムの、またはDNAと言い換えてもいいのですが、DNAの中の遺伝子情報がmRNAに転写されてタンパク質に翻訳される。機能分子のタンパク質が表現系の発現を担っている。こういう直線的な遺伝情報の流れがセントラルドグマだったわけです。 ところがこういうncRNAが大量に発現していて、しかもそのうちのいくつかは機能をもっている。おそらく大半が機能をもっていることが分かっていくと思います。そういうものをわたしたちの細胞は大量に発現している。 そういう場合、従来のセントラルドグマは書き換えられる必要があるだろう。従来のセントラルドグマに加えてRNAによる制御プログラムが存在してそれが複雑で多様な表現系とその変化を規定している。ncRNAはゲノム、あるいはmRNA、あるいはタンパク質に働いて、ゲノムであれば転写の調節、mRNAであれば分解、翻訳の調節、タンパク質とは相互作用して活性のon,offを調節することによって少ない数の遺伝子を使い回すことができるのではないか。つまり、そのかっこよく言えばコンピュータでいうソフトワイヤリングのような役割をncRNAが担っていることによって、限られたセットの重要な因子をいかに使い回すか、っていうことをやっているのだと思います。実際、そうであるかどうかおそらくこれからの研究の非常に重要な方向だと思います。 スライド6 今言ってきたことから見えた重要な問題っていうのは、遺伝子とはどういうものか?ということです。今まではタンパク質をコードするもの、というのと同義だったわけです。ところが、こういうのがわかってきたため、遺伝子というのは次のように書き換わるのではないか。つまり遺伝子とは機能を持ったものを発現する転写ユニットである。それは最終的にタンパク質になるかもしれないし、RNAのままかもしれない。けれども、そこに機能があれば遺伝子と書き換えられるかもしれない。 こういう風に書き換えられるのならば、遺伝子数というのは生物の複雑さと比例しているかもしれない。先程もいいましたように、膨大なncRNAを発現させて機能しているとなると、ncRNAを遺伝子と見なした場合、ヒトの遺伝子は二万二千よりも大きくなって生物の複雑さと相関してくるのではないかと思います。 |
| 立花 | そうすると、今まさに遺伝子の定義が大きく変わるかもしれないというところに来ているのですか? |
| 塩見 | そういうところに来ていると思います。 |
| 立花 | 例えばヒトと線虫は割と変わらないというような見解は一般的でしたが、遺伝子の定義を書き換えなければいけないという認識は分子生物学の世界ではあるのですか?それとも違う意見もあるのですか? |
| 塩見 | 違う意見はあると思います。でそれこそ学生さんに遺伝子の定義とは何かを聞いた場合、面白いと思います。 |
| 志村 | 生物学者一般はそうではないと思いますが、RNAを研究している人たちは一般的にそういう認識をもっていると思います。 |
重要な小さなタンパク質
| 立花 | それでもう一つ質問ですが、ncRNAといった場合、タンパク質をコードするかどうかが切れ目ですが、そのRNAを試料から取り出した場合、そのRNAがコードしているかどうか、それに対応するタンパク質があるかどうかはすぐにはわかりませんよね? |
| 塩見 | わかりません。 |
| 立花 | じゃ、ncRNAかどうかはどうやって判断するのですか? |
| 塩見 | それは非常に重要な問題で、一般的に言われているncRNAの定義は、オープンリーディングフレームを探していって、アミノ酸を100コードできるオープンリーディングフレームが見つからない場合はncRNAと言っています。 だけど、100アミノ酸より短い、例えばペプチドで機能性のものは沢山あります。だから、ncRNAの中には非常に小さい、だけどシグナル伝達とかに関わる重要なタンパク質をコードするようなものもあるわけです。 だから、ncRNAの研究の発展の上に新しい分野が生まれてくるのではないかと思います。例えば、脳の中でも小さな分子作用がコミュニケーションの上で重要であることがわかってきましたようですし、もしかしたら大量に発現しているのかもしれない。それは一つの大きな分野になりますし、今まで隠れていた分野です。 ncRNAの研究はそういった広がりがあります。 それは一番最後のほうに付け加えておいたのですが。そういった研究はイーストのほうでも研究が進められています。例えばパン酵母でも全ゲノムがわかっている。 100アミノ酸以下のオープンリーディングフレームしかとれないものはタンパク質をコードしないという認識で切っていたために、ノンコーディングリージョンて言われていた部位があった。 だけど、本当にコードしないのか?20アミノ酸しか作らないものは何もしていないか?という問いをした人がいまして、一つずつ潰していってミューテーションをとって、網羅的に調べました。すると、ひとつずつ培養する条件を変えてミュータントをとると、だいたい半分は表現形がでるんです。 そういったものは普段は必要ないかもしれないけどある環境の変化に対応するためにもっているのかもしれない。ヒトの場合でも、100以下のアミノ酸しか持たないタンパク質を網羅的に調べていくと重要なことをやっていることが分かっていて、例えばGプロテインのシグナリングに関係しているものは”100アミノ酸以下のタンパク質”というカテゴリーに入ってきます。つまり、シグナル伝達に小さなペプタイドが関係しているのかもしれない。ncRNAの研究の延長上には、そういう、一見ncRNAだが、ノンコーディングではないRNAの新しい機能、小さなペプタイドをコードする機能が見えてくるかもしれません。これは非常に重要で、とくに人間の場合、精神活動に関係しているかもしれない。 |
| 学生A | 厳密な意味で何もコードしないってものはあるのですか? |
| 塩見 | もっと正確にいうと、ただ100以下で切っているんじゃなくて、翻訳されやすいATGで始まっているか、という分け方もあります。 みなさん知っていらっしゃるかもしれませんが、コザックのコンセンサスってのがありまして、開始コドンとして強いATGと弱いATGていうのがあるんです。強いATGコザックのコンセンサスにあてはまるATGは開始コドンとして使われやすい。アミノ酸が100以下で、かつどうみてもコザックのコンセンサスに当てはまらないATGから始まっている。そういう場合は、何もつくっていないんじゃないか、と思われている。 でもそれは実験的に証明されているわけではなくて、進化の過程でたまたま弱いATGが選択されているのかもしれない。普段の時は発現しない。でも何かの時に使うためにわざと弱いやつが転写されているのかもしれない。 いくつかのカテゴリーをいれたプログラムを作り、それに当てはまらないものはncRNAとしています。 |
| 編集者 | 相当恣意的に決めていますよね? |
| 塩見 | そうかもしれません。だから実験的に小さなペプタイドが作られていないか調べなければいけないし、もし作られているとしたら、大きなmRNAの中にたかだか20ぐらいのアミノ酸しかコードしないものがあるということになります。 なぜそういったものがあるか?そしてなぜそういったものが進化の過程で選択されてきたのか?またその翻訳調節がどうやってなされているのか?そういうこともふまえて面白い分野になると思います。 |
RNAの高特異性
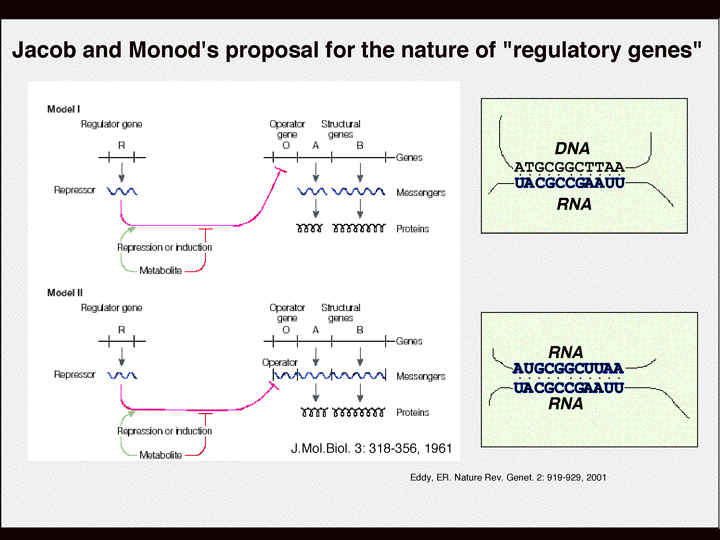
| 塩見 | で、ncRNA研究のもう一つの重要な流れはこうではないかと思います。 今までの研究は、転写やスプライシングやトランスクリプションなどに関わる一般的なファクターの研究だった。 これがここ数年、またはこれからの研究は、個々の遺伝子がどういう風に調節されているか、ということになってきます。つまり最近はやりの言葉でいうと、テーラーメイドの遺伝子発現はどういう風にされているか?個々の遺伝子の選択性はどこからきているのか?例えば、A,B,Cていう遺伝子があるとしてBだけが選択される。その特異性はどこからくるのか?ということです。 スライド7 特異性を出す場合、RNAという分子は非常に都合が良い。 これはジャコブとモノーの非常に有名なオペロン説っていう論文の最後の図です。 スライド8 この最後の図で、遺伝子の発現の調節しているレギュレーター、多分彼らはリプレッサーを想定しているのだと思いますが、それはRNAです、と言っているんですね。でも、このオペロンでは間違いだったのですが、なぜ彼らがRNAだと言っているのかというと、RNAは配列特異性を示す。つまり、塩基の相補性を利用して、特定のDNAまたはRNAを認識できる。しかもRNAで重要なのは、タンパク質と同じように動きまわることができることです。彼らは大腸菌で考えているのですが、標的をさがしながら動き回ることができる。しかも標的を相補性を利用して見つけ出す。こうやってDNAやRNAにくっついて転写の調節や翻訳、RNAの安定性を変えている。 これが1961年の論文なのですが、ぼくらが今やろうとしていることは平たく言うとこういう所なのです。彼らが想定していたリプレッサーっていうのをncRNAだと思えば、ncRNAがDNAに直接作用したり、RNAまたはタンパク質と相互作用することによってゲノム発現をいろいろなレベルでコントロールしている。多くの場合、彼らがリプレッサーと言っているように、ネガティブに調節している、抑制している。そういうのがだんだん見えてきた。 簡単に歴史だけを、二つの論文を使って紹介してきたのですが、ncRNAでもいろいろなタイプがあります。 最近RNAiやmiRNAというのが注目されているように、小分子ncRNAによる遺伝子発現制御機構が注目されたのは、1981年、たぶんこの年が最初の例ではないかと思います。 国立遺伝学研究所の前所長でいらっしゃった富沢純一さんという非常に有名な先生が、コリシンE1というタンパク質の複製を調節するRNAを見つけます。これは直接DNAに作用して、プラスミドと言ってもいいかもしれませんがそれの複製を調節します。1981年はリボザイムの発見の年だったのですが、同じ年にこういう先駆的な発見がなされたわけですね。 この3年後、これは日本人のなのですが、今はニュージャージーにおられる有名な井上さんという方がミックRNAという小さなRNAがその大腸菌のポンプFというタンパク質のmRNAの翻訳を調節することを発見した。 その場合も塩基の相補性を利用して小さなRNAがmRNAと相互作用して負の調節をしています。 おそらくこういう、富沢さん、井上さんの研究がncRNA研究の発端なのではないかと思います。 スライド9 今、大腸菌の話をしましたけど、大腸菌の中で小さなRNAがどういう風に働くか、二つの例を簡単にあげました。一つは、相補性を利用して小さなRNAが標的のmRNAの5‘の開始コドンの少し上流あたりに作用してmRNAの構造を変化させて翻訳を抑制したり促進させたりしています。すなわち、その少し下流にある開始コドンAUGの部分をリボソームがくっつきやすいようにオープンな構造をとったり、あるいは閉じた構造にするかということを、小さなRNA分子が相互作用することで決めています。 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
準備中