レクチャーに使用されたパワーポイントも掲載していますので合わせてご覧ください。
スイッチのON⇔OFF
| 立花 | 相互作用っていうのは具体的にどういうことをやっているのですか? |
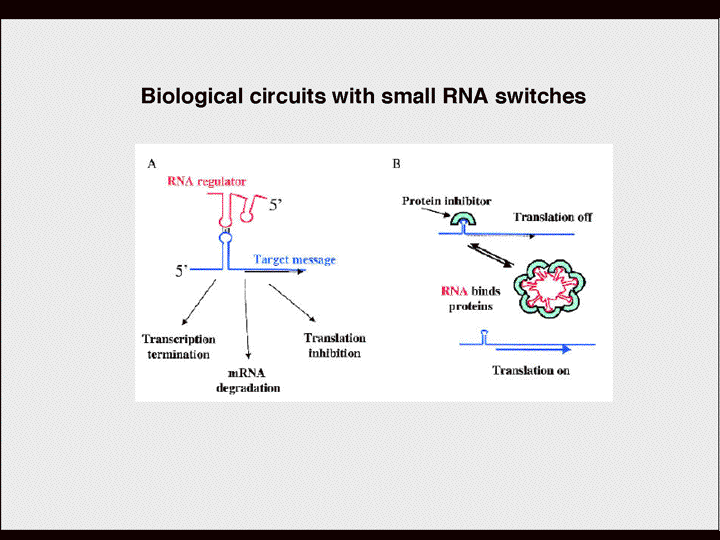
| 塩見 | スライド10 具体的に言いますと、例えばRNAは非常に複雑な二次構造をとっていますよね。開始コドンATGが隠されてしまうような構造をとる時がある。で、大腸菌だとシャルガノっていう配列があって、リボゾームがその配列を認識して最初のATGから翻訳を開始しているのですが、その重要な認識配列がマスクされてしまう構造をとる場合がある。この場合は、翻訳にとってはoffである。 しかし例えばこのRNAと相互作用する小さなRNAがあるとしたら、いままでこういう構造をとっていたのが違った構造をとることによって、オープンな構造になる。水素結合が違う方に取られて、開いた形になった部分をリボゾームが認識して翻訳が開始される。 翻訳がoffからonになるわけです。 その逆もあるわけです。普段はオープンになっててonになってATGあたりは開いていると。だから翻訳はどんどん行われているけど、こういう小さなRNAによって高次構造が変わって、その部分が複雑になってリボソームが入れなくなって翻訳が抑制される。そうして、onからoffになる。 |
| 立花 | 翻訳がon,offされるのはよく分かりましたが、その小さなRNAがそこへくっつくかどうかは何によってわかるんですか?あるいはそのRNAにくっつく他の何かによって進むんですか? |
| 塩見 | わたしたちはRNAが中心なのでRNAがすべてみたいな発言をしていますが、重要なのは転写制御なんです。転写因子による転写制御が重要なんです。 われわれの細胞の中にはさまざまな転写因子があって、それらにはそれぞれ特異性があります。例えば、ABCDというようなシスエレメントがある時、Aというエレメントを認識する転写因子がある一方で、Bというエレメントを認識する転写因子がある。これらの組み合わせが、mRNAやタンパク質の活性を制御する。 転写制御と、こういうレベルでの制御、二つの重要な制御システムがあるんだと思います。mRNAの中にもいろんなシスエレメントが含まれているわけです。 特に、mRNAの場合、5’のUTRと3’のUTRというタンパク質をコードしていない領域がかなりあります。こういう領域には転写因子が認識するシスエレメントが詰まっていて、シスエレメントと転写因子の組み合わせがある。 こういうところを認識する、小さなRNA、つまりncRNA構造というのがおそらくあります。 これによって一つの遺伝子をいろいろなところで制御できる、転写のところでon,offをかけることができる、翻訳のところでもon,offをかけることができて、最終生成物であるタンパク質に対しても活性のon,offをかえることができる。それによって一つの遺伝子を使い回すことができます。それで細胞によって使われるコードが違ってくる。Aという細胞とBという細胞では使われるコードが違う。コードの違いが細胞の違いになっているのかもしれませんが、その組み合わせの複雑さで生物の複雑さができていると思います。限られた遺伝子で複雑な生物ができているのはそのためである、というが最近のコンセプトだと思います。これは単なるコンセプトではなくてシステムバイオロジーによって大量処理するという実験結果によってこういうことがだんだん見えてきています。 それでRNAっていうのは核酸との相補性を利用するだけではなくてタンパク質とも非常に特異性の高い相互作用をします。一つのRNAというのはタンパク質をパートナーにもつことができる。その特異性を利用することによって例えばこういうことができます。 あるmRNAを考えます。そのヘアピン構造の真ん中にある重要なエレメントとよく似たまたは同一のエレメントを多く持つ別のRNAがあるとします。そのエレメントを特異的に認識するタンパク質があります。 このタンパク質が前者のmRNAにひっついている場合は、例えばこの構造を安定させるためにヘアピン構造の中に開始コドンのATGを隠してしまうなどして、その翻訳をoffにします。 ところが、後者のようなRNAがたくさん発現することによって、そちら側にタンパク質がとられ、mRNAが、はじめてオープンな構造になる。他の因子がそこにアクセスすることができる。この場合、他の因子というのはリボソームなどです。 |
| 学生 | Bは具体的にどういう例なんですか? |
| 塩見 | Bは具体的にはトリプトファンなんです。これも大腸菌の例なんですけど、アミノ酸合成でトリプトファンを使う量っていうのは細胞の中で決められているのですが、その量が低下するとトリプトファンの合成する遺伝子をonにする必要があります。その系にこういうものが関わっていることが分かっています。 |
| 志村 | 大分、昔にわかったことです。 |
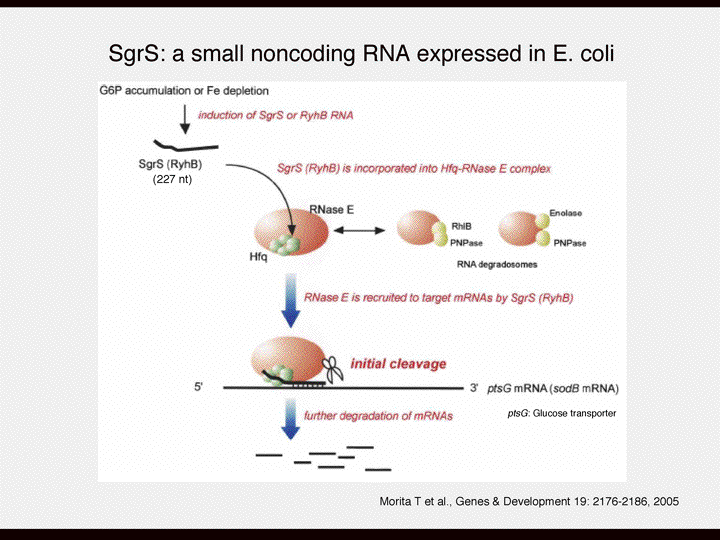
| 塩見 | よくテキストにでてくる例です。 あと最近の例ですが、名古屋大学の理学部の饗場弘二先生のグループの仕事ですけど、彼らがやっているのは糖代謝なんですが、大腸菌のなかのグルコースの量はなぜいつも一定に保たれているのか、というのを研究されていた。 彼らがみつけてきたのは、グルコースの量が増えるとそれをどこかが感知して小さなRNAが作られるようになることです。 細胞表面でグルコースを細胞内に運んでくるタンパク質であるグルコーストランスポーターが、そのmRNAを分解する。そのためにはこのmRNAだけを分解できる特異性が必要である。 グルコーストランスポーターをコードするmRNAと相補性を持つ小さなRNAがあります。小さなRNAがコアとなって、RNAを分解する酵素群と複合体を作る。小さなRNAをガイド分子として、標的mRNAを見つけて分解する。すべてのmRNAを分解するではなくて壊したい、作ってほしくないmRNAだけを分解するようなシステムが大腸菌の中にもある。これも始めの一歩は転写なんです。細胞の中のグルコースの濃度を感知して小さなRNAの発現をonにしてmRNAを分解レベルで制御するわけです。 その組み合わせによって細胞の中のグルコースの濃度が保たれるのです。こういうシステムが出来上がっています。 スライド11 最近の流行りのmiRNAやRNAiといったものは同じ仕組みです。小さなRNAがガイド分子になってRNA分解酵素と複合体を形成してそれが特異性をもって標的mRNAだけを分解して、翻訳を抑制する。特異性をだすために小さなRNAが使われているんです。 |
| 立花 | じゃ、そこらへんは全部特許の対象になるんですか? |
| 志村 | なりますね、将来は。 |
| 塩見 | しかも、良いことに、薬として必要なのはRNAだけで、他のシステムは全部わたしたちの体が持っています。タンパク質とか。しかもRNAはまた代謝されて、RNA合成などの材料になる。だから効果が高い上に、毒性が少ないんです。 |
| 立花 | 人間の体内のホメオスタシスを保っているのはそういった小さな仕掛けが働いているためですか? |
| 塩見 | そうですね、そこらへんはまたあとで話します。 |
転写されるアンチセンス鎖
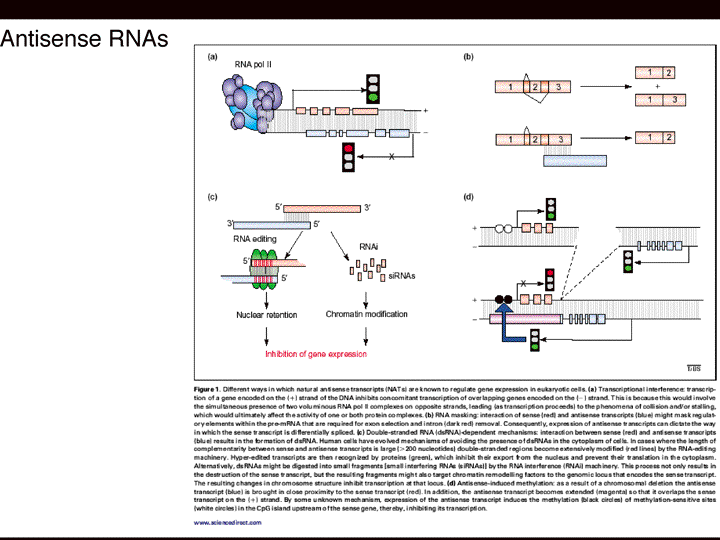
| 塩見 | スライド12 これまで小さなRNAの話をしてきましたが、もう一つ最近重要なのはアンチセンスRNAで、冒頭のところで転写産物の網羅的解析の結果、センスとアンチセンスのペアは非常にたくさんあるということを言いました。 じゃ、反対側の転写される方が何をやっているか?ということになります。たぶんいろいろなことをやっているんだろうけど、最近わかったことをいくつかまとめたものを、ある総説からとってきました。ひとつはセンスが転写されるともう片方は転写されない、つまりこの場合アンチセンスは転写されない。つまり片方がonになるともう片方はoffになる、これは昔からわかっていたことです。 でも最近の転写産物の網羅的解析の結果、こういうのはまれであることが分かってきました。みんな、片方が転写されるともう片方が転写されないと思っていたけど実はそうではない。 じゃ、両方が転写されると何が起こるか?というと、考えられるのは、二つは相補性を持ちますからニ本鎖のRNAになる。 ニ本鎖のRNAがつくられると何がおこるかというと、一つには、核内でニ本鎖のRNAができますと高い頻度で修飾される、Editingされる。例えば、アデニンというのがイノシンになるという修飾を受けます。そういうのがRNAのEditingと言われています。 RNAのEditingが起こるとどうなるかというと、例えばmRNAは核の中で転写されて細胞質にでてきますが、でてこなくなる。mRNAが、核内にとどまっているということは、翻訳に使われないわけですから役に立たないはずなんです。そのレベルでの制御機構があるのかもしれない。もしくは、ポジティブに考えれば、核のなかである役割を果たしてるのかもしれない。つまりRNAのEditingを受けることによってある時は核外にでて翻訳される、ある時は核の中にとどまって新しい役割を果たす。あるいは単純に核から出たり、とどまったりして最終的にできるタンパク質の量を調節しているのかもしれない。これが一つです。 もう一つがRNAiと言われるものです。RNAiっていうのは“ニ本鎖RNAが引き金となって起こる配列特異的な標的mRNAの分解機構”と定義されます。 二本鎖RNAが引き金になるというのは、こういうことです。作られたニ本鎖のRNAを認識してプロセスする酵素があります。プロセスっていうのは分解するんですけど、めちゃくちゃに分解するんじゃなくて22ベースずつぐらいに切っていく酵素がわたしたちの体の中にはあります。そういう酵素によって作られた22ベースぐらいの小さなRNAが先程のように複合体にとりこまれて、最終的に、小さなRNAの相補性を利用することで、標的のmRNAをみつけて分解する。というような機構にも関わっている。 それ以外のいくつかの例ですと、例えば、アンチセンスがセンスと相互作用(アニーリング)することによってスプライシングのパターンを制御している例も見つかっています。 例えばmRNA前駆体から二つの成熟型のmRNAができる場合を考える。エキソン1、2、3が作られる場合と、2をスキップして、エキソン1、3だけが作られる場合とで、二種類のmRNAが作られる。 こういう風にアンチセンスとのアニーリングが行われることによって、片方のスプライシングパターンしか起こらない。 つまり最終的には二種類のタンパク質ができるはずなんですけど、こういうことが働くため一種類のタンパク質しかできない。こういう制御にも関わっている。 |
インプリンティングとncRNA
| 塩見 | もうひとつ、これは非常に重要な分野だと思いますけど、インプリンティングという現象があります。 わたしたちは染色体を対でもっています。片方は父親からもう一方は母親からもらっています。 多くの遺伝子は母親由来のAという遺伝子と父親由来のAという遺伝子が発現しているのですが、ある遺伝子においては母親からの遺伝子しか発現していない、父親からの染色体からはその遺伝子の発現が見られないということがある。 発現していないっていうのは非常に重要で、発現すると病気になるとか、そういうことがわかってきています。 なぜインプリンティングが必要か、いろいろ仮説が出されていますがまだ分かっていない。非常に面白い分野だとおもいます。なぜ、母親由来の、または父親由来の遺伝子しか発現しないのか?また、なぜそのような複雑なシステムが進化の過程で獲得されてきたのか? で、そういうインプリンティングされている遺伝子を調べると、たとえば母親の方だけが発現されている場合、母親の方だけがonになっていて、父親の方はoffになっています。この時、なぜだかしらないけど、父親の方はoffになっている遺伝子の反対側の方はonになっている。 多くの場合、今まで調べられたImprinting geneというのはこういう関係になっています。 つまり、見方を変えて、逆側を遺伝子として捉えると、父親の方はonとなっていまして母親のほうはoffになっています。 面白いことに、片方がタンパク質をコードする遺伝子ならばもう片方がタンパク質をコードしない遺伝子、つまりncRNAをつくります。 そのncRNAが発現することによって、おそらくその反対側のタンパク質をコードする遺伝子の発現を同じ染色体上で抑えているのではないだろうか?こういうことが言われているのですが詳細なメカニズムは分かっていません。 ncRNAはいろんなことをしているということについて、いくつか例をあげました。 インプリンティングや X染色体の不活化にも関与しています。 DNAやRNAに直接結合するのではなくて、タンパク質に直接結合することによってタンパク質の活性を調節している。こういうのをリボレギュレーターといいます。 もう一つは転写抑制といって、上流で読まれることによって、下流の遺伝子発現が抑えられる。上流で読まれることによってポリメラーゼが移動していき、下流の重要な配列に相互作用するような因子と結合するわけです。 こういうところから作られている多くのものはncRNA。つまりその下流の遺伝子のonofをするためだけに転写がon offになっているという遺伝子もどうもありそうだ、という話です。 |
リボレギュレーターの役割
| 塩見 | 今さっき話したimplintingに関わっているようなRNAのリストがあります。 スライド13 リボレギュレーターの二つの例ですけど、一つは東京大学の山本正行先生の有名な仕事です。分裂酵母の減数分裂を制御しているMei2という蛋白質があるんですけど、Mei2は普段は細胞質に存在しているが、減数分裂を進行させるためには核の中に移行しなければならない。 核の中に移行するためにはMeiRNAというnc RNAが発現しなければならない。 減数分裂をするためにはMei2はMeiRNAと複合体を形成することによって、初めて核の中に入っていくことができて、減数分裂を進行させている。 減数分裂という非常に大切な制御は、細胞質から核への移行というレベルで調節されていて、それを制御しているのがRNAであった、ということです。 もうひとつは転写の伸長因子を制御しているPol2。転写の開始点も、伸長過程も制御しています。伸長過程のオンオフを制御している複合体があって、さらに、その複合体のオンオフを制御しているのが小さなRNAであるというのが最近わかってきました。 こういうのがリボレギュレーターと呼ばれています。つまり蛋白質の活性とか、タンパク質の細胞内での局在を制御するようなRNAが存在するんだということです。 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
準備中